ARTÍCULO ORIGINAL
Efectos antibacterianos de extractos de especies del género Ugni que crecen en Chile
Antibacterial effects of extracts of species of the genus Ugni that grow in Chile
Dr. C. Marcia Avello Lorca, Dr. C. Magalis Bittner Berner, Dr. C. José Becerra Allende
Universidad de Concepción. Concepción, Chile.
Introducción: Ugni molinae
Turcz., Ugni candollei (Barnéoud)
O. Berg y Ugni selkirkii (Hook. & Arn.)
Berg son arbustos que se distribuyen en el centro-sur de Chile, incluido el
archipiélago de Juan Fernández. Dada su similitud morfológica
parece que estas especies están estrechamente relacionadas, por lo tanto,
podrían compartir compuestos químicos similares, como sustancias
de tipo fenólico y del tipo terpénico, con reconocida actividad
antimicrobiana, los que han sido identificados en Ugni molinae, la especie
con mayor distribución en Chile.
Objetivo: evaluar la composición química y la actividad antibacteriana
de los extractos de las especies chilenas del género Ugni, y
determinar si existen variaciones entre poblaciones que están creciendo
separadas geográficamente y en diferentes hábitats, como en Chile
continental y el archipiélago de Juan Fernández.
Métodos: se determinó el contenido y la composición
de compuestos químicos por métodos espectrofotométricos
en extractos obtenidos con solventes de polaridad creciente. La actividad antibacteriana
se evaluó a través de inclusión en agar tripticasa frente
a Pseudomonas aeruginosa, Staphyloccoccus aureus y Enterobacter
aerogenes.
Resultados: Ugni candollei y Ugni selkirkii presentan la mayor
concentración de fenoles, flavonoides y taninos totales. Con respecto
a las poblaciones de Ugni molinae, la población del archipiélago
de Juan Fernández presentó la mayor concentración de compuestos
fenólicos y saponinas totales. Si bien existe una tendencia en la actividad
antibacteriana en los extractos metanólicos, que se correlaciona con
la composición química, no se consideran grandes diferencias entre
estas.
Conclusiones: dado que Ugni molinae comparte características
morfológicas con otras especies del género que crecen en Chile,
Ugni candollei y Ugni selkirkii, también comparte características
químicas. Con respecto a las poblaciones de Ugni molinae, se observan
diferencias en el contenido de compuestos fenólicos y saponinas totales,
quizá por las características de hábitat donde estas especies
y poblaciones están creciendo. La mezcla activa de los extractos metanólicos
de especies del género Ugni y las poblaciones de Ugni molinae,
llamada fitocomplejo, concentra las acciones antimicrobianas para las
familias de componentes descritas, sin observarse grandes diferencias entre
estas.
Palabras clave: especies del género Ugni, compuestos fenólicos, terpénicos, actividad antimicrobiana.
Introduction: Ugni molinae Turcz.,
Ugni candollei (Barneoud) O. Berg and Ugni selkirkii (Hook. &
Arn.) Berg are shrubs growing in southern central Chile, including Juan Fernandez
Islands. Given their morphological likeness, these species seem to be closely
related. Therefore, they could share similar chemical compounds, such as phenolic
and terpenic substances with well-known antimicrobial activity. These have been
identified in Ugni molinae, the species with the broadest distribution
in Chile.
Objective: evaluate the chemical composition and antibacterial activity
of extracts of Chilean species of the genus Ugni, and determine the possible
differences between populations growing in geographically separate areas with
dissimilar habitats, such as Continental Chile and Juan Fernandez Islands.
Methods: the content and chemical composition of chemical compounds were
determined by applying spectrophotometric methods to extracts obtained with
increasing polarity solvents. Antibacterial activity against Pseudomonas
aeruginosa, Staphylococcus aureus and Enterobacter aerogenes was
evaluated by inclusion in trypticase agar.
Results: Ugni candollei and Ugni selkirkii exhibit the greatest
concentration of total tannins, phenols and flavonoids. With respect to Ugni
molinae, the populations on Juan Fernandez Islands showed the greatest concentration
of total saponins and phenolic compounds. Although there is a trend in the antibacterial
activity of methanolic extracts that is consistent with their chemical composition,
no great differences are found between them.
Conclusions: Ugni molinae not only shares some morphological features
with other species of the genus growing in Chile Ugni candollei and Ugni
selkirkii. These species also share some chemical characteristics. With
respect to Ugni molinae populations, differences are found in the content
of total saponins and phenolic compounds which are maybe due to the specific
habitat where these species and populations grow. The active mixture of methanolic
extracts of species from the genus Ugni and Ugni molinae populations,
known as phytocomplex, concentrates the antimicrobial activities of the
families of components described above, and no great differences are found between
them.
Key words: species of the genus Ugni, phenolic compounds, terpenic, antimicrobial activity.
INTRODUCCIÓN
El género Ugni Turcz. pertenece a la familia Myrtaceae y comprende entre 19 y 21 taxa que se encuentran desde México, Centro América, Venezuela hasta Chile y Argentina.1 Son arbustos o árboles pequeños de hoja perenne que se cultivan tanto por su importancia ornamental como por el atractivo de sus frutos. De estas, 3 se encuentran en Chile: Ugni molinae Turcz., Ugni candollei (Barnéoud) O. Berg y Ugni selkirkii (Hook. & Arn.) Berg; son arbustos que se distribuyen en el centro-sur de Chile incluido el archipiélago de Juan Fernández.
Aunque no existen estudios evolutivos del grupo, dada su similitud morfológica pareciera que estas especies están estrechamente relacionadas. Debido a esta relación podrían compartir compuestos químicos similares. Solo se tiene conocimiento de la composición química de U. molinae, especie que ha sido evaluada con respecto a su actividad biológica.2-5 Estos autores han descrito la presencia de sustancias de tipo fenólico como ácidos fenólicos y flavonoides, y del tipo terpénico como ácidos triterpénicos pentacíclicos compuestos con reconocida actividad antimicrobiana.6,7 La amplia distribución de esta especie podría afectar la composición química de las poblaciones de una región a otra. Se tiene antecedentes que factores geoclimáticos como la radiación solar, la humedad y la fertilidad del suelo pueden afectar la síntesis de metabolitos secundarios,8 así también como factores bióticos.9 U. molinae está presente en el archipiélago de Juan Fernández como una especie introducida. Este archipiélago está a más de 600 km de la costa chilena y posee características de clima y suelo bastante particulares, las que podrían estar afectando la composición química de las poblaciones de esta especie que allí crecen.
Dado que U. molinae comparte características morfológicas con otras especies del género que crecen en Chile, U. candollei y U. selkirkii, quizá compartan características químicas y consecuentemente desarrollen actividad biológica similar, por lo tanto, el objetivo de este trabajo era evaluar la composición química y la actividad antibacteriana de los extractos de las especies chilenas del género Ugni, así como determinar si existen variaciones entre poblaciones que están creciendo separadas geográficamente y en diferentes hábitats, como en Chile continental y el archipiélago de Juan Fernández.
MÉTODOS
Material vegetal
El material biológico de las especies nativas del género Ugni se recolectó en época de floración (noviembre- marzo 2008-2009): U. candollei, Región de Los Ríos (Valdivia, Parque Oncol, cerro Oncol); U. selkirkii, archipiélago de Juan Fernández (Isla Robinson Crusoe, Mirador Selkirk, cerro Portezuelo); U. molinae, Región del Bío-Bío (Barrio Universitario, Concepción), y las poblaciones de la Región de la Araucanía y en el archipiélago de Juan Fernández, Isla Robinson Crusoe. Las especies fueron identificadas por el taxónomo Dr. Roberto Rodríguez, Facultad de Ciencias Naturales y Oceanográficas, Universidad de Concepción. Un ejemplar de cada especie quedó registrada en el Herbario CONC (125465, 162345, 146511, 151285 y 116887, respectivamente). Para la comparación de las especies del género Ugni se consideró en forma arbitraria la población de U. molinae de la región del Bío-Bío como representante de esta especie.
Obtención de extractos
El material biológico se deshidrató a la sombra, a temperatura ambiente y se redujo de tamaño en molino de cuchillas. Para la obtención de extractos se procesaron 50 g de las muestras molidas en aparato Soxhlet. Se utilizaron en forma sucesiva los solventes siguientes: hexano, cloroformo, acetato de etilo, y metanol, hasta agotamiento del material vegetal. La relación masa solvente fue de 1:6. Cada extracto se concentró en rotoevaporador y se llevó a sequedad en liofilizador. Se realizaron reacciones de reconocimiento de metabolitos secundarios en cada extracto (reacción de Shinoda para el reconocimiento de flavonoides, reacción con FeCl3 en taninos y la prueba de espuma en el reconocimiento de saponinas). Para las determinaciones químicas y biológicas se seleccionaron los extractos obtenidos con acetato de etilo y metanol por su contenido en compuestos fenólicos y saponinas.
Cuantificación de metabolitos secundarios
Fenoles totales: se determinó mediante el método de Folin-Ciocalteu. La determinación de fenoles totales se expresó como equivalentes de ácido gálico por gramo de planta seca (g EAG/g materia seca).10
Flavonoides totales: se realizó según Salamanca y otros.11 Las determinaciones se expresaron como equivalentes de quercetina por gramo de planta seca (g EQ/g materia seca).
Taninos totales: se realizó según describe Lastra y otros,12 y se expresó como equivalentes de ácido gálico por gramo de planta seca (g EAG/g materia seca).
Saponinas totales: la determinación se efectuó como describe Hideaki y otros.13 Los resultados se expresaron como gramos de saponina por gramos de planta seca (g saponina/g materia seca).
Actividad antibacteriana
Se utilizaron cepas bacterianas aisladas desde productos patológicos del cepario del Departamento de Microbiología, Facultad de Ciencias Biológicas de la Universidad de Concepción, las que fueron Pseudomonas aeruginosa (ATTC 27653), Staphyloccoccus aureus (ATTC 6538P) y Enterobacter aerogenes (UC-1). La actividad antibacteriana se determinó de acuerdo con la metodología descrita por Liu y otros.14 Para esto, las cepas se diluyeron y así obtener una densidad celular aproximada de 1x106 ufc/mL, luego se sembraron por inclusión en agar tripticasa. En estos cultivos en agar, se construyeron pocillos de 7 mm de diámetro, en los cuales se depositaron alícuotas de 100 µL de los extractos (1 mg/mL) en suero fisiológico. Las placas se incubaron a 37 ºC y se observó la posible presencia de halos de inhibición a las 24 h. Como blanco se utilizó suero fisiológico y amoxicilina (25 µg/mL) como estándar.
RESULTADOS
Cuantificación de metabolitos secundarios
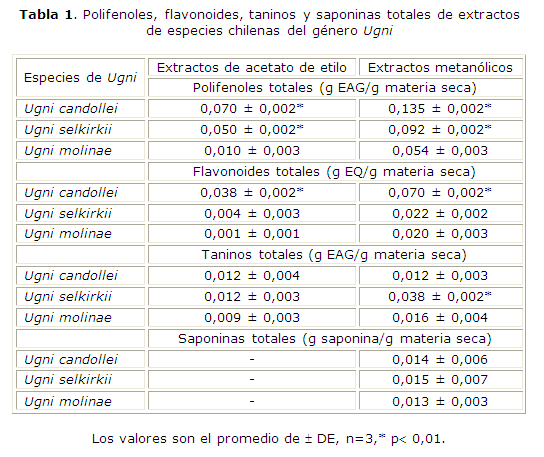
Se cuantificaron polifenoles totales en los extractos obtenidos con acetato de etilo y metanol, de las especies y poblaciones en estudio, donde se observó la presencia de flavonoides y taninos por las reacciones de reconocimiento cualitativas. Como se observa en la tabla 1, U. candollei y U. selkirkii son especies con alta concentración de polifenoles totales, tanto en los extractos obtenidos con acetato de etilo como con metanol. En este último resultaron mayores, lo que indica la naturaleza polar de los compuestos involucrados. U. candollei presenta los mayores niveles incluidos los flavonoides totales (0,038 ± 0,002 g EQ/g materia seca en el extracto acetato de etilo y 0,070 ± 0,002 g EQ/g materia seca en el extracto metanólico), mientras que U. selkirkii destaca en el contenido de taninos totales en el extracto metanólico (0,038 ± 0,002 g EAG/g materia seca). Con respecto al contenido de saponinas totales se cuantificaron en los extractos metanólicos, en los cuales se observó la presencia de saponinas por las pruebas de reconocimiento cualitativas. No se observaron diferencias significativas entre especies, alcanzando un máximo de 0,015 g saponinas/g materia seca en U. selkirkii.
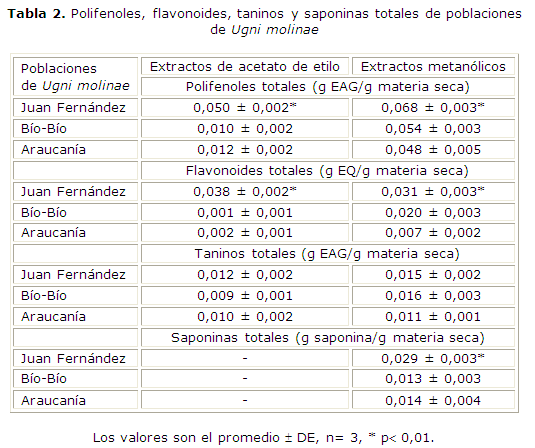
Con respecto a las poblaciones de U. molinae (tabla 2), la población del archipiélago de Juan Fernández es la que contiene la más alta concentración de polifenoles totales, tanto en el extracto obtenido con acetato de etilo (0,050 ± 0,002 g EAG/g materia seca) como con metanol (0,068 ± 0,003 g EAG/g materia seca). Esta población también muestra los mayores niveles de flavonoides totales en ambos extractos (0,038 ± 0,002 g EQ/g materia seca en el extracto acetato de etilo y 0,031 ± 0,003 g EQ/ g materia seca en el extracto metanólico). Las poblaciones continentales, en cambio, presentan menores niveles de polifenoles totales en comparación con la insular, aun así son considerables en los extractos metanólicos (0,054 ± 0,003 g EAG/g materia seca en la población de la región del Bío-Bío y 0,048 ± 0,005 g EAG/g materia seca en la población de la región de la Araucanía). Entre ambas poblaciones el contenido de flavonoides totales es mayor en la población de la región del Bío-Bío (0,020 ± 0,003 g EQ/g materia seca). Con respecto al contenido de taninos totales no existen diferencias significativas entre las tres poblaciones, concentrándose en los extractos metanólicos. Con respecto al contenido de saponinas totales se cuantificaron en los extractos metanólicos, en los cuales se observó la presencia de saponinas por las pruebas de reconocimiento cualitativas. La población del archipiélago de Juan Fernández es la que presenta el mayor contenido de saponinas totales (0,029 ± 0,003 saponina/g materia seca) .
Actividad antibacteriana
Los extractos acetato de etilo y metanólicos fueron estudiados frente a bacterias patógenas humanas. Los resultados que a continuación se presentan corresponden al diámetro de halo de inhibición de crecimiento bacteriano observados en presencia de ambos tipos de extractos. Si bien los resultados (de los extractos metanólicos) se muestran muy interesantes, no fue posible determinar la concentración mínima inhibitoria (CMI) debido a la interacción de los propios componentes de los extractos con el contenido del medio de cultivo.
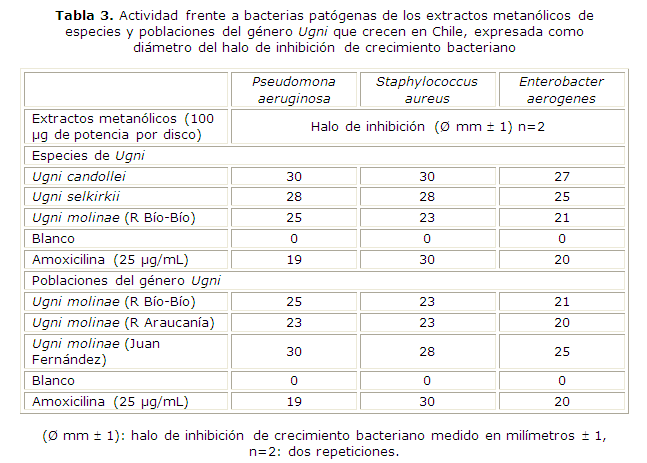
Como se observa en la tabla 3, la especie que produjo mayores halos de inhibición en las tres cepas bacterianas fue U. candollei en su extracto metanólico, sigue en efectividad el extracto metanólico de U. selkirkii y U. molinae.
Estos resultados coinciden con el contenido de fenoles y saponinas totales determinados con el espectrofotómetro en los extractos estudiados (coeficiente de correlación igual a 0,837, p< 0,05).
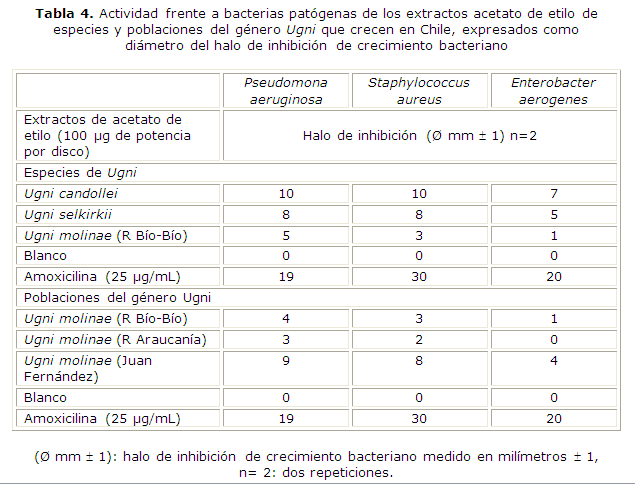
Los resultados de los halos de inhibición de los extractos obtenidos con acetato de etilo se observan en la tabla 4. Tanto para las especies del género Ugni, como para las poblaciones de U. molinae, los halos de inhibición representan baja actividad frente a las cepas bacterianas.
DISCUSIÓN
Los resultados proponen una importante interacción entre los componentes de los extractos metanólicos de las especies y poblaciones estudiadas con compartimentos bacterianos que condicionan la supervivencia de estos microorganismos. Las diferencias entre los resultados de los extractos metanólicos y los obtenidos con acetato de etilo, se deben a la respectiva composición química cualitativa y cuantitativa. Es de esperar encontrar compuestos de polaridad intermedia en los extractos de acetato de etilo y una concentración más baja de compuestos polihidroxilados y glicosilados, con relación a los extractos obtenidos con metanol, que serían componentes con acción perturbadora en las barreras defensivas bacterianas.15-17
Es interesante hacer una relación con respecto al lugar de recolección de las especies del género Ugni y poblaciones de U. molinae, y la composición química y consecuente actividad biológica. Todas las especies fueron recolectadas sobre el nivel del mar, en suelos volcánicos y cercanos al mar. La especie U. candollei fue recolectada en el cerro Oncol a 715 m sobre el nivel del mar. El cerro pertenece al Parque Oncol, que es de conservación, privado, ubicado en la comuna de Valdivia, región de Los Ríos a 29 km de la ciudad (http://www.parqueoncol.cl). Esta especie es la que concentra los niveles más altos de polifenoles y flavonoides; factores que podrían contribuir a este fenómeno lo constituyen la exposición a la radiación, la alta humedad ambiental en el sector y la influencia de las constantes precipitaciones. Se ha descrito la participación directa de estos factores en la biosíntesis de metabolitos fenólicos según Treutter, Wu y otros, así como Harbaum-Piayda y otros.18-20 De igual forma la especie U. selkirkiii fue recolectada en la cima del Mirador Selkirk, ubicado en el cerro Portezuelo a 565 m sobre el nivel del mar, en el archipiélago de Juan Fernández (http://www.comunajuanfernandez.cl). Mientras que la especie U. molinae se recolectó en los valles de la región del Bío-Bío y también la población de la Región de la Araucanía.
La especie recolectada en el archipiélago de Juan Fernández, U. selkirkii, debe sus características químicas, probablemente, al clima por la alta humedad y el origen volcánico del lugar.8 El alto contenido en oxihidróxidos de hierro y aluminio, con deficiencia en el aporte de nitrógeno, fósforo, potasio y azufre favorece la biosíntesis tanto de compuestos fenólicos como de terpenoides, entre estos las saponinas.8,21,22 Lo que diferencia los suelos de la isla con los del continente, también de origen volcánico, es que estos últimos son especiales a nivel de superficie y cercanía a los cráteres, lo cual explicaría niveles más importantes de nutrientes y la escases de otros.
La humedad, las características del suelo y la radiación, podrían influir en las características genéticas que determinan las características fenotípicas de las especies, porque la respuesta adaptativa a las condiciones de su hábitat natural, hace que las funciones de estos compuestos se vuelvan más importantes como lo describen Waniska, Vivanco y otros, Davies y Schwinn, así como Vermerris y otros.23-26 Los factores bióticos también deberían influir en las características químicas de las especies vegetales, pero para las especies de Ugni en Chile no existen estudios en este sentido. De esta forma podrían interpretarse las diferencias cuantitativas entre especies.
La población de U. molinae del archipiélago de Juan Fernández se considera una especie invasora y plaga en el territorio insular, una de las posibles razones de su éxito podría constituirlo la importante composición de metabolitos secundarios. La alta humedad, los suelos de origen volcánico del archipiélago y la erosión podrían influir favorablemente en la biosíntesis de compuestos de naturaleza terpénica, como las saponinas, y de compuestos fenólicos en una especie introducida8.
Con respecto a la actividad frente a patógenos humanos, los extractos metanólicos de las especies y poblaciones en estudio resultaron los activos. La especie que produjo mayores halos de inhibición en las cepas bacterianas estudiadas fue U. candollei, sigue en efectividad U. selkirkii, y U. molinae. En cuanto a las poblaciones de la especie U. molinae, los mayores halos de inhibición se observaron con la población del archipiélago de Juan Fernández. Estos resultados coinciden con el contenido de fenoles y saponinas totales determinado mediante espectrofotómetro.
Alberto y otros, Shan y otros, así como Rodríguez y otros,15-17 hacen evidente una mayor inhibición de los extractos de polifenoles en bacterias grampositivas que en bacterias gramnegativas, debido a la sensibilidad de las (+) frente a especias y extractos de plantas. Esta mayor sensibilidad en el caso de Staphylococcus aureus puede ser atribuida a la estructura de su membrana citoplasmática, la cual permite que las sustancias antibacterianas puedan fácilmente destruir la membrana de la célula y resultar en una salida del citoplasma.
Los compuestos fenólicos, en general, actúan por diversos mecanismos dependiendo de su estructura; interaccionando con receptores, enzimas y formando uniones con compuestos como proteínas.27 Así, los compuestos fenólicos antibacterianos por excelencia son los taninos, como los derivados gálicos. Debido a su peso molecular, conformación química y proporción en la mezcla vegetal, a través de formación de enlaces con los grupos amida de proteínas e interacciones hidrofóbicas causan alteraciones e inviabilidad celular.28 Los mecanismos que lo explican incluyen precipitación de proteínas y el efecto prooxidante.29 Esto último ocurre cuando en la oxidación de los taninos se producen altos niveles de radicales semiquinonas y quinonas; la autooxidación fenólica ocurre en la presencia de oxígeno y la dismutación en semiquinonas formando quinonas. La activación redox de hierro, por ejemplo, podría potenciar la oxidación de los taninos, cuando los niveles de oxígeno son limitados y las quinonas pueden formar altos niveles de especies reactivas del oxígeno.
El extracto metanólico de U. selkirkii destaca en el análisis cuantitativo por su contenido en taninos totales. Los taninos presentes en los extractos estudiados mantienen niveles importantes en las demás especies y poblaciones estudiadas. En los extractos metanólicos de las especies U. candollei y U. selkirkii y en el de la población U. molinae del archipiélago de Juan Fernández se reporta la presencia de ácidos fenólicos. Los ácidos orgánicos provocan una reducción en el pH extracelular en microorganismos, aumentando el turgor celular que provoca un incremento de la presión mecánica sobre la pared de estos, lo cual hace que eventualmente estalle.28
Los terpenos, en general, cumplen una función antimicrobiana fundamental dada su naturaleza lipofílica. Los grandes terpenos han mostrado interacción con barreras lipídicas de microorganismos desestabilizando la integridad de las membranas, como la superficie lipofílica de la membrana externa de las gramnegativas, la cual actúa como una barrera que limita la entrada de agentes antibacterianos. Esta membrana es rica en moléculas de lipopolisacáridos y está asociada con las enzimas en el espacio periplasmático las cuales son capaces de romper las moléculas introducidas desde el exterior.
Las saponinas interaccionan con barreras lipofílicas por afinidad química de la genina, y la porción glicosídica contribuye a la disrupción celular. Pero la totalidad de estos mecanismos no ha sido del todo descrita y no se ha podido establecer la relación estructura-actividad.30 Se ha propuesto la formación espontánea de complejos entre moléculas de saponinas y colesterol de membrana, que conducirían a la formación de poros. Estos poros causarían un incremento en la permeabilidad de la membrana, permitiendo la circulación de iones y macromoléculas entre proteínas, además de la extracción de esteroles a través de vesículas. Estos fenómenos pueden proporcionar la explicación más simple de los cambios observados en la conductividad de iones y la movilidad de proteínas a través de membranas.
Existen otros modelos que siguen los mismos pasos descritos antes, pero con algunas variantes, como la interacción de saponinas con regiones ricas en esfingomielina y colesterol.30 En consecuencia, la formación de estos complejos da como resultado la separación de fases en las membranas. Las especies U. candollei, U. selkirkii, y U. molinae continentales mostraron niveles importantes de glicósidos de ácidos triterpénicos. La población de U. molinae del archipiélago de Juan Fernández superó ese contenido. Aún así, no se observan diferencias significativas entre especies y poblaciones que puedan estar indicando la acción preferente de compuestos fenólicos o terpénicos.
Dado que U. molinae comparte características morfológicas con otras especies del género que crecen en Chile, U. candollei y U. selkirkii, comparten también características químicas, por lo cual constituyen las especies con mayor concentración de fenoles, flavonoides y taninos totales. Con respecto a las poblaciones de U. molinae, la población del archipiélago de Juan Fernández presentó la mayor concentración de compuestos fenólicos y saponinas totales, quizá por las características de hábitat donde estas especies y poblaciones están creciendo. La mezcla activa de los extractos metanólicos de especies del género Ugni y poblaciones de U. molinae llamada fitocomplejo concentra las acciones antimicrobianas para las familias de componentes descritas. Si bien, existe una tendencia en la actividad antibacteriana que se correlaciona con la composición química, no se consideran grandes diferencias entre estas.
AGRADECIMIENTOS
Este trabajo fue apoyado por CONICYT 24091006; Dirección de Posgrado, Universidad de Concepción, D.I. 210.074.043.
REFERENCIAS BIBLIOGRÁFICAS
1. Mabberley DJ. The plant book. England: Cambridge University Press; 1997. p. 80.
2. Aguirre MC, Delporte C, Backhouse N, Erazo S, Letelier ME, Cassels BK, et al. Topical anti-inflammatory activity of 2 á-hydroxy pentacyclic triterpene acids from the leaves of Ugni molinae. Biorganic Medicinal Chemistry. 2006;14:5673-7.
3. Rubilar M, Pinelo M, Ihl M, Scheuermann E, Sineiro J, Nuñez MJ. Murta leaves (Ugni molinae Turcz.) as a source of antioxidant polyphenols. J Agricultural Food Chemistry. 2006;54:59-64.
4. Delporte C, Backhouse N, Inostroza V, Aguirre MC, Peredo N, Silva X, et al. Analgesic activity of Ugni molinae (murtilla) in mice models of acute pain. J Ethnopharmacol. 2007;112:162-5.
5. Rubilar M, Jara C, Poo Y, Acevedo F, Gutierrez C, Sineiro J, et al. Extracts of Maqui (Aristotelia chilensis) and Murta (Ugni molinae Turcz): sources of antioxidant compounds and ß-glucosidase/-ß-amylase inhibitors. J Agricultural Food Chemistry. 2011;59:1630-7.
6. Avello M, Valdivia R, Mondaca MA, Ordóñez JL, Bittner M, Becerra J. Actividad de Ugni molinae Turcz. frente a microorganismos de importancia clínica. Boletín Latinoamericano y del Caribe de Plantas Medicinales. 2009;8(2):141-4.
7. Avello M, Valdivia R, Sanzana R, Mondaca MA, Mennickent S, Aeschlimann V, et al. Extractos a partir de berries nativos para su uso como preservantes naturales en productos cosméticos. Boletín Latinoamericano y del Caribe de Plantas Medicinales. 2009;8(6):479-86.
8. Oh M, Trick H, Rajashekar C. Secondary metabolism and antioxidants are involved in environmental adaptation and stress tolerance in lettuce. J Plant Physiology. 2009;166:180-91.
9. Charudattan R, Dinoor A. Biological control of weeds using plant pathogens: accomplishments and limitations. Crop Protection. 2000;19:691-5.
10. Velioglu Y, Mazza G, Gao L, Oomah B. Antioxidant activity and total phenolics in selected fruits, vegetables and grain products. J Agricultural Food Chemistry. 1998;46:4113-7.
11. Salamanca G, Correa I, Principal J. Perfil de flavonoides e índices de oxidación de algunos propóleos colombianos. Zootecnia Tropical. 2007;(25)2:95-102.
12. Lastra H, Rodríguez E, Ponce H, González ML. Método analítico para la cuantificación de taninos en el extracto acuoso de romerillo. Rev Cubana Plant Med. 2000;5(1):17-22.
13. Hideaki O, Shigeo K, Shoji S. Separation and determination of saponins of bupleuri radix by droplet counter current chromatography CDCC. Planta Medica. 1978;33:152-9.
14. Liu D, Kwasniewska K, Chau Y, Dutkta B. A tour-hour agar plate for rapid toxicity assessment of water-soluble and water-insoluble chemical. Environmental Toxicology Waters Quality. 1991;6:437-44.
15. Alberto M, Rinsdahl M, Manca M. Antimicrobial effect of polyphenols from apple skins on human bacterial pathogens. Environmental Biotechnol J. 2006;9(3):1-5.
16. Shan B, Cai Y, Brooks J, Corke H. The in vitro antibacterial activity of dietary spice and medicinal herb extracts. International J Food Microbiology. 2007;117(1):112-9.
17. Rodríguez M, Tomassini L, Manca M, Strasser A. Antioxidant capacity and antibacterial activity of phenolic compounds from Argentinean herbs infusions. Food Control. 2009;21(5):779-85.
18. Treutter D. Significance of flavonoids in plant resistance and enhancement of their biosynthesis. Plant Biology. 2005;7:581-91.
19. Wu JY, Wong K, Ho KP, Zhou LG. Enhancement of saponin production in Panax ginseng cell culture by osmotic stress and nutrient feeding. Enzyme Microbial Technology. 2005;36:133-8.
20. Harbaum-Piayda B, Walter B, Bengtsson G, Hubbermann EM, Bilger W, Schwarz K. Influence of pre-harvest UV-B irradiation and normal or controlled atmosphere storage on flavonoid and hydroxycinnamic acid contents of pak choi (Brassica campestris L. spp. chinensis var. communis). Postharvest Biology Technology. 2010;56:202-8.
21. Kovacik J, Klejdus B, Backor M, Repcak M. Phenylalanine ammonia-lyase activity and phenolic compounds accumulation in nitrogen-deficient Matricaria chamomilla leaf rosettes. Plant Science. 2007;172:393-9.
22. Giorgi A, Mingozzi M, Madeo M, Speranza G, Cocucci M. Effect of nitrogen starvation on the phenolic metabolism and antioxidant properties of yarrow (Achillea collina Becker ex Rchb.). Food Chemistry. 2009;114:204-11.
23. Waniska RD. Structure, phenolic compounds and antifungal proteins of Sorghum caryotipes. In: Chandrashekar A, Bandyopadhyay R, Hall AJ, editors. Proceedings of International Consultation. India: ICRISTAT; 2000. p. 72.
24. Vivanco J, Cosio E, Loyola V, Flores H. Mecanismos químicos de defensa en las plantas. Investigación y Ciencia. 2005;341:68-75.
25. Davies K, Schwinn K. Molecular biology and biotechnology of flavonoid biosynthesis. In: Flavonoids Chemistry, Biochemistry and Applications. USA: Taylor & Francis Group. LLC.; 2006. p. 1151-2..
26. Vermerris W, Nicholson R. Phenolic compound biochemistry. The Netherlands: Springer-Dordrecht; 2006. p.1-285.
27. Larcher W. Physiological plant ecology. Austria: Springer-Verlag; 2003. p. 1-517.
28. Barbehenn R, Constabel C. Tannins in plant-herbivore interactions. Phytochemistry. 2011;72:1551-65.
29. McArt SH, Spalinger DE, Collins WB, Schoen ER, Stevenson T, Bucho M. Summer dietary nitrogen availability as a potential bottom-up constraint on moose in south-central Alaska. Ecology. 2009;90:1400-11.
30. Augustin J, Kuzina V, Andersen S, Bak S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry. 2011;72:435-57.
Recibido: 18 de noviembre de 2012.
Aprobado: 07 de enero de 2013.
Marcia Avello Lorca. Casilla 237. FONO: 56-041-2204523; FAX: 56-041-2207086. Concepción, Chile. Correo electrónico: maavello@udec.cl
{kind=link}
{kind=link}