ARTÍCULO ORIGINAL
Aislamiento del estigmasterol de las semillas de Crotalaria juncea L. (cascabelito) y su bioactividad sobre Drosophila melanogaster
Isolation of stigmasterol from seeds of Crotalaria juncea L. (sunn hemp) and bioactivity against Drosophila melanogaster
Giann Carlos Peñaloza Atuesta,I Carlos Alberto Peláez JaramilloII
I
Universidad del Tolima, Ibagué, Colombia.
II Universidad
de Antioquia, Medellín, Colombia.
RESUMEN
Introducción:
En el campo de los productos naturales, las investigaciones que procuran la
purificación e identificación de los principios activos de las plantas
junto con la determinación de sus actividades biológicas se consolidan
como una línea relevante del desarrollo científico.
Objetivos:
Evaluar la actividad biológica de la fracción de éter de petróleo
de las semillas de Crotalaria juncea L. sobre Drosophila melanogaster,
así como identificar y caracterizar estructuralmente los principios bioactivos.
Métodos:
Se evaluó la bioactividad del extracto de éter de petróleo (fracción
45-60 °C) obtenido de las semillas secas de C. juncea ingerido por
D. melanogaster. La estructura química del compuesto se determinó
con diferentes pruebas de resonancia magnética nuclear (RMN).
Resultados:
La bioactividad del compuesto se manifiesta como la inhibición de los estados
larvarios, porque disminuye el número de pupas de los tratamientos. La
relación dosis/respuesta permitió calcular una CI50 de
4,24 × 10-4 M para esta bioactividad. Los resultados de diferentes
pruebas de RMN, 1-D 13C, 1-D 1H y 2-D 1H-13C
HMBC, HMQC, COSY y la comparación con la literatura permitieron definir
la estructura del compuesto como estigmasterol.
Conclusiones:
El ensayo de la bioactividad del compuesto aislado en el extracto de polaridad
baja permitió comprobar actividad insecticida promisoria, ya que disminuye
el número de pupas de los tratamientos comparado con los controles. El
análisis estructural de este compuesto permitió determinar que se
trata del estigmasterol y realizar el primer informe
de su aislamiento en las semillas de C. juncea. Anteriormente se pudo
constatar que D. melanogaster no puede metabolizar el estigmasterol.
Palabras clave: Crotalaria juncea L.; correlación dosis/respuesta; Drosophila melanogaster; estigmasterol; modelo biológico; relación adulto/pupa (R A/P), CI50.
ABSTRACT
Introduction:
In the field of natural products research, studies aimed at purification
and identification of active principles in plants and determination of their
biological activities have become a relevant, consolidated line of scientific
development.
Objectives:
Evaluate the biological activity of the petroleum ether fraction of Crotalaria
juncea L. seeds against Drosophila melanogaster, and identify and
structurally characterizethe its bioactive principles.
Methods:
Evaluation was conducted of the bioactivity of petroleum ether extract (45 to
60 ºC fraction) obtained from C. juncea dry seeds after ingestion
by D. melanogaster. The chemical structure of the compound was determined
with several different nuclear magnetic resonance (NMR) tests.
Results:
Bioactivity of the compound manifests as inhibition of larval stages, since
a reduction occurs in the number of pupas under treatment. The dose-response
relationship allowed estimation of a IC50 of 4.24 × 10-4
M for this bioactivity. Results of the different NMR tests, 1-D 13C, 1-D 1H
and 2-D 1H-13C HMBC, HMQC, COSY, and comparison with the literature made it
possible to define the structure of the compound as stigmasterol.
Conclusions:
Bioactivity testing of the compound isolated from the low polarity extract revealed
promising insecticidal activity due to a reduction in the number of pupas under
treatment as compared to controls. Structural analysis of the compound led to
its identification as stigmasterol and a first report was written about its
isolation from C. juncea seeds. It had been previously verified that
D. melanogaster does not have the ability to metabolize stigmasterol.
Key words: Crotalaria juncea L., dose-response relationship, Drosophila melanogaster; stigmasterol, biological model, adult/pupa ratio (APR), IC50.
INTRODUCCIÓN
Crotalaria juncea L., también conocida como Sunn hemp, pertenece a la familia Fabaceae, considerada la familia más grande con alredor de 600 especies.1 C. juncea ha sido utilizada en la India para la producción de fibras. En sistemas agrícolas se ha probado su capacidad alelopática en las semillas de maíz.2 Como cultivo intermedio C. juncea ha demostrado controlar el nemátodo Meloidogyne incognit,3 además de ser usada como abono verde por su capacidad fijadora de nitrógeno.4
El género Crotalaria es la principal fuente de alcaloides pirrolizidínicos (PAs, por sus siglas en inglés) de metabolitos con un amplio efecto toxicológico que va desde el daño hepático en humanos y mamíferos hasta la aparición de cáncer en los animales sometidos a altas dosis de estos compuestos.5 Por otro lado, se ha reportado el uso de trampas a base de PAs como atrayentes químicos para reducir el daño causado por Zonocerus variegatu.6 Además de los PAs, existe una amplia lista de metabolitos secundarios aislados de numerosos miembros de esta especie entre los que se pueden encontrar aminas, flavonoides, cumarinas, lectinas y fitoesteroles.1,7
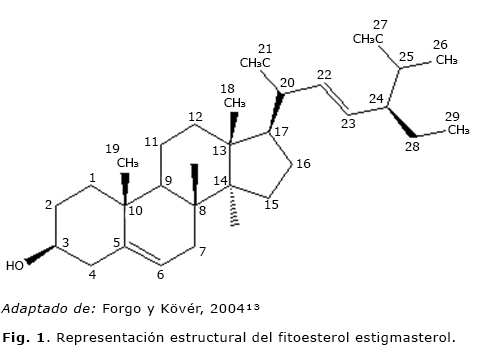
En este trabajo se informa sobre el aislamiento y la elucidación estructural de un fitoesterol (Fig. 1) conocido y reportado en otras especies del género, pero aquí se obtiene por primera vez de las semillas de C. juncea.
Los esteroles forman parte de los componentes lipídicos de una dieta y constituyen la mayor proporción de la fracción no saponificable de los lípidos.8,9 En los aceites y en las grasas vegetales existe una gran variedad de fitoesteroles, los más comunes son el sitoesterol, el campesterol y el estigmasterol.8,10 Se ha demostrado que los fitoesteroles como el β-sitoesterol reducen la producción de tumores en animales tratados con carcinógenos de colon,9 particularmente el estigmasterol ha demostrado tener un notable efecto antiinflamatorio al inhibir varios mediadores favorecedores de la inflamación, actividad que se atribuye, en parte, a la inhibición de la ruta NF-κB en condrocitos humanos y de ratones, lo que le ha conferido un importante potencial contra la osteoartritis.11 En la figura 1 se muestra la representación estructural del estigmasterol aislado y el sistema de numeración de la molécula está basado en la regla de la biosíntesis de la cadena lateral.12
A diferencia de otros organismos, los insectos deben obtener los esteroles de su dieta, debido a su imposibilidad para sintetizarlos de novo.14,15 En estos organismos los esteroles juegan dos papeles primordiales, uno de ellos es no metabólico, constituido por los esteroles que forman parte de las membranas (esteroles estructurales) y por otra parte se encuentran los esteroles metabólicos12 que se convierten en precursores de hormonas esteroidales capaces de regular las diferentes etapas del desarrollo, por ejemplo, la 20-OH-ecdisona que regula los procesos de muda.10
En muchas especies de insectos, incluido los dípteros, el esterol predominante es el colesterol, ya que es el precursor necesario para la síntesis de la ecdisona u hormona de la muda,10,12,14-16 pero en las plantas el contenido de colesterol es bajo, lo que significa que los insectos fitófagos deben ser capaces de metabolizar los fitoesteroles hasta colesterol para garantizar los contenidos mínimos que aseguren la supervivencia de los organismos.13 Se ha verificado que en las especies de orugas como Manduca sexta y en los dípteros como Aedes aegypti, el metabolismo de los fitoesteroles actúa por vía de la desalquilación sobre el carbono C24.10,15 En Drosophila melanogaster se ha confirmado su capacidad para convertir ergosterol en colesterol, pero también su incapacidad para metabolizar sitoesterol o desmoesterol hasta colesterol.15
El objetivo de este trabajo es evaluar la actividad biológica sobre D. melanogaster de la fracción de éter de petróleo de semillas de C. juncea, así como identificar y caracterizar estructuralmente sus principios bioactivos.
MÉTODOS
Colonias de Drosophila melanogaster (cepa Canton)
La colonia de Drosophila melanogaster (Canton), genéticamente estable, se obtuvo en el laboratorio del Grupo Interdisciplinario de Estudios Moleculares (GIEM) del Instituto de Química de la Universidad de Antioquia, donde se mantuvo a una temperatura promedio de 23 °C con una humedad relativa de 65 % y un periodo de luz de 12 h por más de 700 generaciones.
Material vegetal
Las semillas de Crotalaria juncea L. (Familia Fabaceae) fueron suministradas por el Centro Internacional de Agricultura Tropical (CIAT) de Palmira, Valle del Cauca, Colombia. El material se encuentra depositado en el programa de recursos genéticos bajo el número 21 709 en el municipio ubicado en la región sur del Valle del Cauca (Colombia) a una altura de 1 001 m.s.n.m y con una temperatura promedio de 27 °C.
Preparación del material vegetal
Las semillas se obtuvieron secas, se molieron en un molino marca CONDUX-WERK® modelo LV15M, el material homogenizado se extrajo con éter de petróleo (fracción 45-60 °C) mediante el método Soxhlet. El extracto se concentró por rotaevaporación asistida al vacío en un equipo marca BÜCHI® modelo RE 111. El compuesto purificado se emulsificó utilizando el tensoactivo TWEEN-80 (Polioxietilen sorbitan monooleato). Para garantizar su solubilidad en el alimento de D. melanogaster se adicionó 7 % (v/v) del tensoactivo a una mezcla agua/estigmasterol y la mezcla se sometió a la acción mecánica para disminuir el tamaño de partícula y homogenizarla.
Bioensayo: interrupción del ciclo de vida
Las pruebas de bioactividad dirigida17 se realizaron en frascos de vidrio transparente de 5 cm de ancho × 10 cm de alto con una capacidad aproximada de 50 mL. Para evaluar el ciclo de vida de D. melanogaster, el modelo biológico se sometió a la ingestión del alimento mezclado con el compuesto emulsificado. Para este fin se depositó en los viales el volumen de emulsión correspondiente a cada concentración a evaluar (0,0 - 3,1 × 10-3 M) y se completó hasta un volumen final de 15 mL con el alimento para D. melanogaster que se preparó con 450 mL de agua, 7 g de agar, 50 g de harina de maíz, 15 g de levadura, 40 mL de melaza y 2,5 mL de ácido propiónico. Los viales se dejaron en reposo por 12 h antes de realizar el cruce de los individuos.
Los individuos adultos y jóvenes de 3 días de edad de D. melanogaster se expusieron al alimento durante 48 h en una proporción macho:hembra 3:3, tiempo después del cual se descartó la vía parental para realizar el recuento de pupas y de adultos a los 8 y 14 días, respectivamente.
Aislamiento y purificación de los metabolitos
La fracción de desengrase (éter de petróleo, fracción 45-60 °C) se concentró en un rotoevaporador. En este proceso se precipitó un sólido cristalino. Para purificar este sólido se retiró el aceite utilizando medios físicos; el remanente se sometió a extracción con hexano grado HPLC. En este solvente los cristales no se solubilizaron y se limpiaron para posteriormente recristalizarse en cloroformo.
Identificación de los metabolitos
Se identificó el principio activo purificado mediante espectroscopia de resonancia magnética nuclear en un equipo marca BRUKER® AMX 300 con 300 MHz para 1H y 75 MHz para 13C. Se realizaron las pruebas 1-D 13C, 1-D1H y se tomaron los espectros en cloroformo deuterado (CDCl3) a 25 °C empleando tetrametil silano (TMS) como estándar interno.
Análisis estadístico
Todos los experimentos se realizaron con tres repeticiones independientes (n= 3). Se efectuó el análisis de regresión con el método de mínimos cuadrados para las variables número de individuos y los diferentes niveles de concentración (M) del compuesto purificado. Se obtuvo un modelo ajustado y se analizaron los datos atípicos teniendo en cuenta los residuos estudentizados mayores que 2. Se compararon las medias de los tratamientos mediante un ANOVA entre ensayos con el paquete estadístico Statgraphics Centurión XV. Los análisis se realizaron con un valor de significancia del 0,05, la evaluación de la normalidad de los datos se realizó con la prueba Shapiro-Wilk.
RESULTADOS
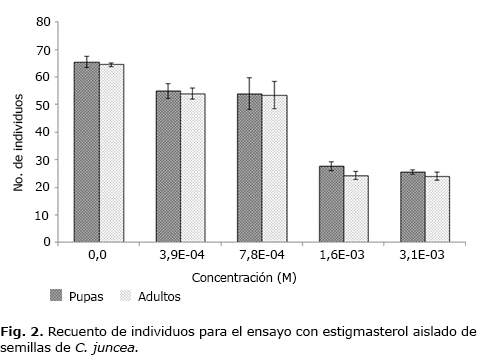
La figura 2 muestra el recuento en los estados pupa y adulto del modelo D. melanogaster sometido a la ingestión de alimento con suplemento de estigmasterol emulsificado.
El recuento de los individuos pupa y adulto permitió establecer que el
estigmasterol no presenta actividad biológica durante la transformación
de pupa a adulto como se puede apreciar en la figura 2,
pero es evidente que existe disminución en el número de pupas de los
tratamientos con respecto a los controles no tratados. En la figura
2 se aprecia también que la relación adulto/pupa permanece constante
en todo el intervalo de concentraciones. Se propuso un modelo matemático
que describe la relación entre las variables concentración y respuesta:
con alto grado de significancia estadística, al 95 % de confianza, ya que
el análisis de varianza presenta un p< 0,05 (0,0334) y no se
encuentran residuos atípicos. Con el modelo de correlación se calculó
una CI50 de 4,24 × 10-4 M para la actividad inhibitoria
del estigmasterol sobre los estados inmaduros (larvas) del modelo biológico.
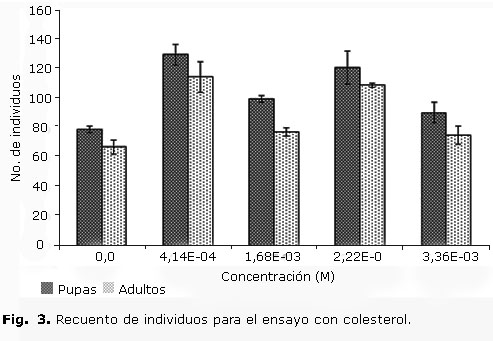
El comportamiento de D. melanogaster frente al estigmasterol hace pensar que en esta especie el mencionado compuesto no sigue la ruta bioquímica para su conversión en colesterol. Con el objetivo de valorar, al menos preliminarmente, la respuesta de D. melanogaster frente al colesterol, se realizaron bioensayos utilizando colesterol estándar (Cholest-5-en-3β-ol; Art. 3670 MERCK®). Es necesario aclarar que en el caso de la dieta con suplemento de estigmasterol no se eliminó el colesterol endógeno del medio de prueba que puede provenir de las sustancias con las que se elaboró el alimento (levadura, agar, melaza, entre otros.). En la figura 3 se muestran los resultados de este ensayo.
Hay que resaltar que la relación adulto/pupa para todo el intervalo de concentraciones evaluado también se mantiene aproximadamente constante. En esta prueba con suplemento de colesterol es importante señalar que en el intervalo de concentraciones evaluado el análisis estadístico de las variables no permitió establecer una correlación significativa entre ellas, ya que p> 0,05 evidenció un aumento del número de individuos de los tratamientos, lo que explica la actividad inhibitoria del estigmasterol sobre D. melanogaster.
Análisis estructural del metabolito purificado
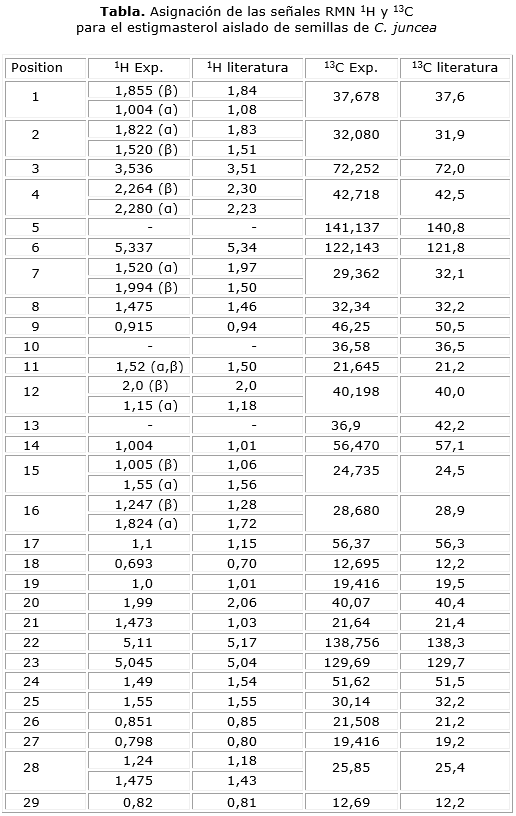
En la tabla se muestra la asignación de las señales de 1H RMN y 13C RMN, la cual se realizó utilizando la combinación de diferentes pruebas de RMN 1-D 13C, 1-D1H2-D 1H-13C HMBC, 1H-13C HMQC, 1H-1H COSY con lo que se logró proponer la estructura química del estigmasterol como el compuesto responsable de la actividad observada.
La gran complejidad de los espectros, particularmente la de los espectros acoplados, dificulta el análisis estructural de los esteroles. Sobre la base de las evidencias experimentales obtenidas en las diferentes pruebas de RMN, así como de la comparación con los reportes de la literatura (tabla), se dedujo que la estructura del compuesto aislado en la fracción de éter de petróleo de semillas de C. juncea coresponde al estigmasterol por la similitud de los desplazamientos químicos.
DISCUSIÓN
Los esteroles son sustancias fundamentales para el desarrollo de los insectos, ya que estos no pueden sintetizarlos de novo, contrariamente a lo que ocurre en los mamíferos.14 En estos organismos el colesterol es el precursor de las hormonas esteroidales como la ecdisona (hormona de la muda) y muchos de ellos lo obtienen mediante el metabolismo de los fitoesteroles ingeridos en su dieta. En los insectos como D. melanogaster, la biosíntesis de ecdisona necesita que el colesterol consumido en la dieta sea desaturado a 7-dehidrocolesterol en las glándulas protorácicas, principalmente, donde se conoce que la reacción es catalizada por el citocromo P450.15
En ese sentido, aunque el comportamiento de los individuos de D. melanogaster en diferentes estadios carece de significancia estadística en la prueba con suplemento de colesterol estándar, es importante señalar que, para todos los tratamientos del estudio, el número de pupas y adultos es mayor que para el control no tratado, lo que indica que posiblemente D. melanogaster posee la habilidad de incorporar de su dieta este colesterol estándar y utilizarlo para incrementar su capacidad de desarrollo.
El análisis de la bioactividad del estigmasterol sobre D. melanogaster permitió establecer, al menos al prinicpio, que el insecto no posee la habilidad para metabolizar y utilizar este fitoesterol en su normal desarrollo, del mismo modo que se ha demostrado para otros fitoesteroles como desmoesterol, sitoesterol, colestanol,16,18 sin importar la presencia de colesterol endógeno en el alimento.
El estigmasterol inhibe el desarrollo de las etapas larvarias de D. melanogaster, actividad evidenciada con una concentración de 1,6 × 10-3 M con la cual el número de pupas disminuyó el 42,3 % comparado con el control no tratado. Por el contrario, cuando se comparó el blanco con una concentración de colesterol estándar de 1,68 × 10-3 M se evidenció que el número de pupas aumentó en un 20,69 % con respecto al control no tratado. Este tipo de actividad del estigmasterol sobre el desarrollo larvario también ha sido evaluado en larvas de Heliothis zea. En este insecto la tasa de desarrollo de las larvas que consumieron estigmasterol en su dieta resulta comparable con el crecimiento de larvas con dietas con suplememento de colesterol,13 efecto contrario al observado en este trabajo sobre D. melanogaster.
Es importante resaltar que en los dos experimentos que se comparan el número de individuos presentes en los controles no tratados es aproximadamente igual, lo que indica la estabilidad del modelo biológico utilizado en las condiciones del análisis. También es importante mencionar que existe una inhibición del paso pupa-adulto inherente al modelo que produce valores de esta relación inferiores a 1 para los controles.
Mediante el análisis de los modelos de regresión fue posible demostrar que el estigmasterol purificado de semillas de C. juncea presentó actividad inhibitoria promisoria sobre D. melanogaster, actividad que puede darse en alguno de los estadios larvarios o sobre la viabilidad de los huevos. En futuras investigaciones se determinará el estadio en el cual se afecta D. melanogaster.
En el análisis estructural del estigmasterol, particularmente en el espectro 1H-RMN, se probó una serie de señales complejas entre δ 1,0 y 2,0 ppm causadas por el solapamiento de los protones metilénicos (CH2) y metínicos (CH), señales que son características en los espectros 1H-RMN de los esteroles.
Dentro de las señales de 1H-RMN se destacan cuatro que se desplazan a campo alto, δ 5,35, δ 5,15, δ 5,05 y δ 3,5 ppm que corresponden a protones ubicados sobre los carbonos 6, 22, 23 y 3, respectivamente (Fig. 1). El protón sobre C6 hace parte del esqueleto principal del esterol y en el espectro acoplado protón-protón se puede verificar que existe acoplamiento con el protón 7β y un acoplamiento a larga distancia con los protones sobre C4 (Hα y Hβ), el protón en C3 también hace parte del esqueleto del esterol y está enlazado al carbono oximetínico (CH-OH). En el espectro acoplado protón-protón se pueden verificar los acoplamientos con los protones ubicados sobre los carbonos C2 (Hα y Hβ) y con los protones sobre C4 (Hα y Hβ). Los protones sobre C22 y C23 hacen parte de la cadena lateral del esterol y corresponden a protones vinílicos. En el espectro acoplado protón-protón se identifican los acoplamientos existentes entre estos protones y los protones en C20 y C24, respectivamente.
En el espectro 13C-RMN se verifica la presencia de señales correspondientes a 29 carbonos, dentro de ellas las asignadas a dos carbonos olefínicos (C23) δ 129,69 y (C22) δ 138,756 ppm, y adicionalmente el carbono oximetínico (C3) δ 72,252 ppm. En el experimento DEPT135 se diferencian las señales correspondientes a seis grupos metil (CH3), nueve grupos metileno (CH2), once grupos metino (CH) y tres carbonos cuaternarios. Los desplazamientos químicos de 1H-RMN y 13C-RMN para el compuesto aislado de C. juncea y los espectros bidimensionales concuerdan con los reportados en la literatura;19 por lo tanto, es posible proponer la estructura química del estigmasterol como el compuesto responsable de la actividad inhibitoria sobre la aparición de pupas de D. melanogaster.
En resumen, la prueba de bioactividad del compuesto aislado en el extracto de polaridad baja permitió comprobar la actividad insecticida promisoria que disminuye el número de pupas de los tratamientos en comparación con los controles. El análisis estructural de este compuesto definió que se trata del estigmasterol y permitió informar por primera vez su aislamiento en semillas de C. juncea. Además, se pudo establecer el precedente de que D. melanogaster no tiene la habilidad de metabolizar el estigmasterol.
REFERENCIAS BIBLIOGRÁFICAS
1. Devendra BN, Srinivas N, Solmon KS. A comparative pharmacological and phytochemical analysis of in vivo & in vitro propagated Crotalaria species. Asian Pac J Trop Med. 2012;5(1):37-41.
2. Cruz-Silva CTA, Matiazzo EB, Pacheco FP, Nóbrega LHP. Allelopathy of Crotalaria juncea L. aqueous extracts on germination and initial development of maize. Idesia. 2015;33(1):27-32.
3. Curto G, Dallavalle E, Santi R, Casadei N, D'Avino L, Lazzeri L. The potential of Crotalaria juncea L. as a summer green manure crop in comparison to Brassicaceae catch crops for management of Meloidogyne incognita in the Mediterranean area. Eur J Plant Pathol. 2015;142:829-41.
4. Hubbard RK, Strickland TC, Phatak S. Effects of cover crop systems on soil physical properties and carbon/nitrogen relationships in the coastal plain of southeastern USA. Soil Tillage Res. 2013;126:276-83.
5. Xia Q, Yan J, Chou MW, Fu PP. Formation of DHP-derived DNA adducts from metabolic activation of the prototype heliotridine-type pyrrolizidine alkaloid, heliotrine. Toxicol Lett. 2008;178(2):77-82.
6. Timbilla JA, Lawson BW, Yeboah-Gyan K. Field Evaluation of a Novel Pyrrolizidine Alkaloid Attracticide for the Management of Zonocerus variegatus on Cabbage. J Appl Sci. 2007;7(4):575-9.
7. Wink M, Mohamed GIA. Evolution of chemical defense traits in the Leguminosae: mapping of distribution patterns of secondary metabolites on a molecular phylogeny inferred from nucleotide sequences of the rbcL gene. Biochem Syst Ecol. 2003;31(8):897-917.
8. Careri M, Elviri L, Mangia A. Liquid chromatography-UV determination and liquid chromatography-atmospheric pressure chemical ionization mass spectrometric characterization of sitosterol and stigmasterol in soybean oil. J Chromatogr A. 2001;935(1-2):249-57.
9. Calkins BM, Whittaker DJ, Nair PP, Rider AA, Turjman N. Diet, nutrition intake, and metabolism in populations at high and low risk for colon cancer. Am J Clin Nutr. 1984;40:896-905.
10. Jing X, Grebenok RJ, Behmer ST. Sterol / steroid metabolism and absorption in a generalist and specialist caterpillar: Effects of dietary sterol / steroid structure, mixture and ratio. Insect Biochem Mol Biol. 2013;43(7):580-7.
11. Gabay O, Sanchez C, Salvat C, Chevy F, Breton M, Nourissat G, et al. Stigmasterol: a phytosterol with potential anti-osteoarthritic properties. Osteoarthr Cartil. 2010;18(1):106-16.
12. Nes WD, Zhou W, Guo D, Dowd PF, Norton RA. Sterol utilization and metabolism by Heliothis zea. Lipids. 1997;32(12):1317-23.
13. Forgo P, Kövér KE. Gradient enhanced selective experiments in the 1H NMR chemical shift assignment of the skeleton and side-chain resonances of stigmasterol, a phytosterol derivative. Steroids. 2004;69(1):43-50.
14. Behmer ST, Nes WD. Insect Sterol Nutrition and Physiology: A Global Overview. In: Advances in Insect Physiology. Acad Press Ltd, London. 2003;31(3):1-72.
15. Jing X, Vogel H, Grebenok RJ, Zhu-salzman K, Behmer ST. Dietary sterols / steroids and the generalist caterpillar Helicoverpa zea: Physiology, biochemistry and midgut gene expression. Insect Biochem Mol Biol. 2012;42(11):835-45.
16. Feldlaufer MF, Weirich GF, Imberski RB, Svoboda JA. Ecdysteroid production in Drosophila melanogaster reared on defined diets. Insect Biochem Mol Biol. 1995;25(6):709-12.
17. Warren JT, Bachmann JS, Dai J, Gilbert LI. Differential Incorporation of Cholesterol and Cholesterol Derivatives into Ecdysteroids by the Larval Ring Glands and Adult Ovaries of Drosophila melanogaster: a Putative Explanation for the l (3) ecd1 Mutation. Insect Biochem Mol Biol. 1996;26(8):931-43.
18. Kircher W, Gray MANN. Cholestanol-cholesterol utilization by axenic Drosophila melanogaster. J Insect Physiol. 1978;24(6-7):555-9.
19. Peñaloza GC, Peláez CA. Evaluación de la actividad biológica de extractos de Crotalaria juncea mediante el modelo Drosophila melanogaster. VITAE. 2008;15(2):279-84.
Recibido:15 de
julio de 2016.
Aprobado: 4 de marzo
de 2017.
Giann Carlos
Penaloza Atuesta. Universidad del Tolima, Ibagué, Colombia.
Correo electrónico:
gcpenalozaa@ut.edu.co